
Metabolismo ruminale e metanogenesi
• 30 Luglio 2021

Il rumine può essere considerato un fermentatore popolato da miliardi di microrganismi, responsabili delle trasformazioni della sostanza organica proveniente dalla dieta. La comunità microbica nel suo complesso comprende i batteri, gli archea, i protozoi ed i funghi. Ciascuno di questi nell’ecologia ruminale svolge un ruolo preciso, fondamentale e complesso che sta alla base dei processi digestivi. I batteri e gli archea, nonostante siano entrambi organismi monocellulari e procarioti, sono differenti ed appartengono a due domini distinti. Morfologicamente, gli archea si distinguono dai batteri perché la loro membrana cellulare è costituita da eteri del glicerolo o del glicerolo fosfato, con catene isopreniche (alifatiche ramificate) mentre quelle batteriche sono costituite da esteri di fosfoglicerolo, con acidi grassi saturi o insaturi. Inoltre, nelle membrane degli archea non è presente l’acido muramico principale costituente del peptidoglicano, componente esclusivo della parete cellulare batterica, alla quale conferisce rigidità. Le principali differenze, però, si trovano a livello della trascrizione e traduzione del patrimonio genetico. Da questo punto di vista gli archea sono più simili agli eucarioti che non ai batteri stessi. Questi microrganismi, infatti, hanno tratti di r-RNA molto simili agli eucarioti e le loro polimerasi molto più complesse di quelle presenti nelle cellule batteriche e, a livello del DNA, presentano degli introni, ovvero delle sequenze di basi azotate non codificanti, come negli organismi più evoluti. Da un punto di vista strettamente metabolico i batteri si distinguono dagli archea perché non producono metano mentre questa via metabolica è propria degli archea.
I rapporti fra i microrganismi ruminali sono estremamente complessi e i vari gruppi microbici hanno tra loro relazioni di mutualismo o antagonismo. Inoltre, sono fortemente influenzati dalla dieta con particolare riferimento alla quantità e qualità della componente fibrosa. Molte delle attività dei microrganismi ruminali sono sinergiche, ovvero l’attività di alcuni di essi dipende dai metaboliti di altri. Un esempio è proprio la metanogenesi, che è operata dagli archea che riciclano l’idrogeno (H2) prodotto durante l’acetogenesi condotta dai cellulosolitici.
Attualmente sono state caratterizzate circa 70 specie di archea di cui 21 sono le più importanti. Fra di esse ricordiamo il Methanobacterium formicicum, il Methanobacterium bryantii, il Methanobrevibacter ruminantium, il Methanobrevibacter millerae, il Methanobrevibacter olleyae, il Methanomicrobium mobile, il Methanoculleus oletangyi.
Nelle diete per ruminanti, i carboidrati rappresentano la principale fonte energetica e sono contenuti principalmente nella componente foraggiera della razione; il circa 70-80% della razione fornisce l’energia necessaria sia all’animale ospite sia ai microrganismi. Grazie proprio alla presenza dei microrganismi del prestomaco, i ruminanti riescono ad ottenere energia anche dai carboidrati strutturali come la fibra, cosa che non è possibile per un monogastrico. Durante la degradazione anaerobica della componente fibrosa viene prodotta una grande quantità di H2, che a sua volta è trasformato dagli archea in metano (CH4) come una sorta di difesa, poiché il suo accumulo è nocivo per il microbiota. In questa maniera, una volta neutralizzato viene eliminato mediante successiva eruttazione dell’animale.
Fino a qualche anno fa, la perdita di metano attraverso l’eruttazione veniva considerata come una mera «perdita» energetica della razione, decretandone l’efficienza alimentare: tanto più è elevato il tasso della metanogenesi, tanto meno è efficiente la dieta. Oggi, invece, la produzione di metano da parte dei ruminanti viene considerata anche a livello globale come un problema di tipo ambientale, in quanto il metano è un gas serra (green-house gas, GHG), che concorre insieme al nitrossido (N2O) e all’anidride carbonica (CO2) ai danni all’atmosfera, con conseguenti cambiamenti climatici e surriscaldamento della Terra. Si stima che l’allevamento concorra per il 19% all’emissione totale di gas a effetto serra.
Il principale costituente della fibra è la cellulosa, a sua volta formata da glucosio che viene degradato a piruvato con la glicolisi. Il bilancio di queste reazioni cataboliche, che dal glucosio portano alla formazione di acetato, è tale che per ogni mole di glucosio si forma una mole di CH4. Il pH ottimale per la produzione di CH4 è 7.0-7.2 ma si può formare anche nell’intervallo 6.6-7.6, anche se a questi valori l’attività fibrolitica (che è quella che favorisce la via dell’acetato) è più limitata.
Idrogeno e anidride carbonica sono la base per la sintesi del metano, grazie all’attività dei batteri metanogeni (Metanogenesi enterica) i quali, comunque, possono utilizzare anche altri substrati: formiato, acetato, metanolo, metilammine, metil-solfuri che sono intermedi di reazione nella via per la metanogenesi. Queste ultime vie alternative sono, però, energeticamente più sfavorite a causa di un ∆G meno. Pertanto, se vogliamo diminuire la produzione di metano è necessario incentivare le vie metaboliche ruminali che sottraggono l’H2. Alcuni microrganismi ruminali possono competere con i metanogeni, sottraendo H2. Questi sono i batteri acetogenici. Questa via metabolica è stata osservata in diversi erbivori ed è particolarmente sviluppata in quelli non ruminanti per “smaltire” l’accumulo di H2 derivante dalla fermentazione intestinale della fibra.
Nei ruminanti è stato osservato che il numero dei batteri acetogenici è elevato nei primi mesi di vita ma poi diminuisce con l’età a favore di un incremento dei metanogeni. Ciò è dovuto a competizione. Nell’adulto gli acetogeni sono in numero circa dieci volte minore rispetto ai metanogeni, anche se tali differenze possono variare in funzione della dieta animale.
Normalmente la metanogenesi è termodinamicamente favorita rispetto all’acetogenesi, poiché in condizioni normali di funzionamento del rumine il ΔG della prima è circa il doppio in valore assoluto rispetto a quello della seconda (−135 kJ vs. −72 kJ). Questa sintesi, inoltre, è anche cineticamente favorita poiché la concentrazione minima di H2 a cui si innescano le reazioni per i metanogeni è circa 10-100 volte inferiore rispetto a quella necessaria per gli acetogeni. Questa è la ragione per la quale la metanogenesi enterica è fortemente favorita. Anche le bioidrogenazioniconcorrono a consumare H2 ma sono poco efficienti!
L’equilibrio fra acidi grassi volatili (AGV), formati a livello ruminale, è lo specchio delle vie metaboliche attivate dai microrganismi durante la fermentazione dei carboidrati. Pertanto, per riuscire a stimare le emissioni di metano è necessario avere un dato certo sulla produzione di AGV in funzione della dieta. Esistono vari modelli meccanicistici che permettono di predire la produzione di AGV, ma il problema più grosso è riuscire a stimare il tasso di assorbimento reale dell’animale a livello delle pareti ruminali. Normalmente le equazioni inseriscono un coefficiente di correzione calcolato mediante studi che hanno messo a confronto il dato analitico con quello rilevato attraverso l’uso di camere respiratorie.
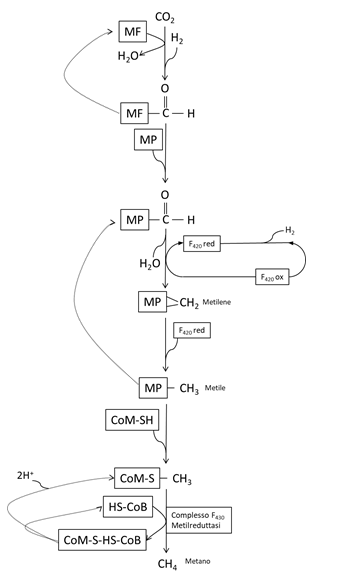
Figura 1 – Le principali vie biochimiche della metanogenesi nei ruminanti.
F430,F420 coenzimi essenziali alla metanogenesi
MF, metanofurano
MP, metanopterina
CoM, coenzima M
CoB, coenzima B
Articolo di Arianna Buccioni e Federica Mannelli – Dipartimento di Scienze e Tecnologie Agrarie, Alimentari, Ambientali e Forestali – Università degli Studi di Firenze
Foto: Pixabay
emissioni metabolismo ruminanti
Last modified: 30 Luglio 2021









